上月的文献学习,我们梳理了一篇影响因子7分左右的关于乙酰化的SCI,文章涉及乙酰化研究的实验不多,是不是意犹未尽呢?今天,我们一起来看看影响因子16左右的文章怎么研究乙酰化的吧~
这次分享的文献题目是:KAT6A Acetylation of SMAD3 Regulates Myeloid-Derived Suppressor Cell Recruitment, Metastasis, and Immunotherapy in Triple-Negative Breast Cancer,影响因子为16.803分,两个月前才热乎乎的发表在Adv Sci杂志上。
想了解高分文章是怎么研究乙酰化的伙伴们,现在就跟着我,踏着之前文献解读的“标准步骤”往下看吧~
一、KAT6A的过表达与三阴性乳腺癌(TNBC)转移相关
为了确定KAT6A在乳腺癌转移中的作用,作者首先用METABRIC dataset 分析了16个赖氨酸乙酰转移酶(KAT)基因DNA拷贝数的改变情况。结果显示,KAT基因在TNBC中表现出不同程度的拷贝数扩增,而KAT6A拷贝数扩增最多(Figure 1A)。Kaplan-Meier分析发现KAT6A扩增拷贝数高的TNBC患者,总生存期更差(Figure 1B)。此外,高表达的KAT6A亦与无复发生存率的降低显著相关(Figure 1C)。接下来的免疫组化结果显示远处转移的TNBC肿瘤KAT6A表达明显高于无肿瘤转移的TNBC(Figure 1C-1D)。与原发性TNBC肿瘤相比,KAT6A在淋巴结和肺转移中均显著上调(Figure 1F)。细胞水平检测发现,与MCF-10A细胞相比,KAT6A在所有检测的乳腺癌细胞中都有高表达,具有高转移能力的TNBC细胞中KAT6A蛋白水平高于非转移细胞(Figure 1G)。在MDA-MB-231和BT-549细胞中敲低KAT6A(Figure 1H)显著降低了细胞的增殖能力(补充材料),侵袭和迁移能力(Figure 1I - 1J)以及原位异种移植瘤的生长和转移能力(Figure 1K-1L)。同时,KAT6A的敲低亦可显著抑制转移性肺结节的形成(Figure 1M- 1N)、延长动物的生存期(Figure 1O)。这些数据表明,KAT6A对TNBC转移至关重要,KAT6A高表达的患者预后较差。二、KAT6A使SMAD3在Lys20和Lys117发生乙酰化
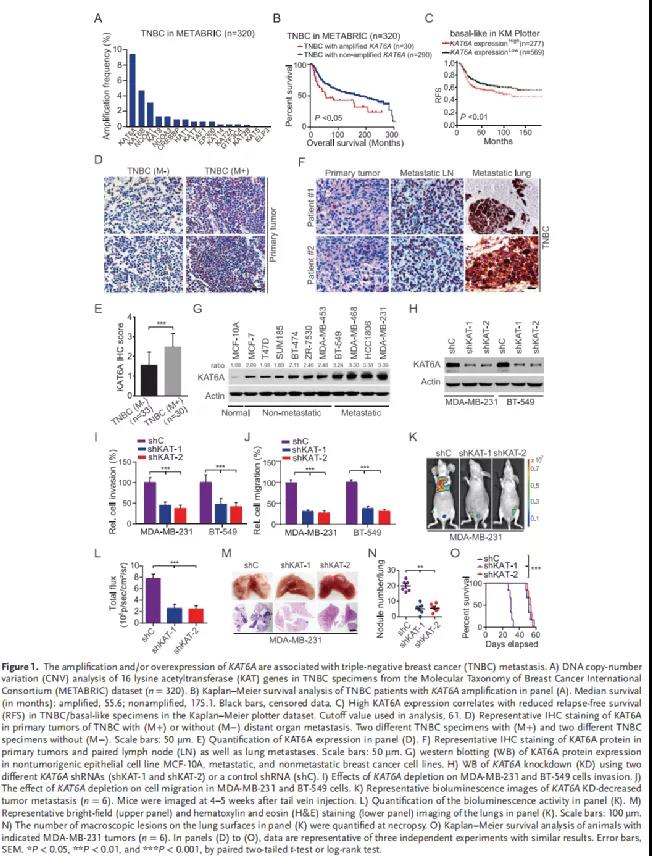
为了探索KAT6A在乳腺癌转移中调控的下游靶点,作者从MDA-MB-231细胞中纯化了flag标记的KAT6A复合物,并进行MS分析。在鉴定出KAT6A可结合蛋白中,SMAD3高度富集(Figure 2A)。外源性以及内源性Co-IP分析均证实KAT6A与SMAD3的存在相互作用(Figure 2B-2C)。GST pulldown实验发现纯化的重组KAT6A可直接与SMAD3相互作用,TGF-β1的刺激显著增强KAT6A-SMAD3的关联(Figure 2D)。在HEK293T细胞中,KAT6A与野生型SMAD3或缺失突变体SMAD3共表达分析发现:SMAD3的c端MH2结构域(氨基酸232-425)是与KAT6A关联所必需的(Figure 2E-2F)。接下来,作者评估了KAT6A 敲低对SMAD3乙酰化的影响。TGF-β1激活SMAD3的乙酰化(Figure 2G),而KAT6A的敲低减弱了TGF-β1诱导的SMAD3乙酰化(Figure 2H)。与野生型 KAT6A相比,KAT6A乙酰转移酶活性缺陷突变体G657E或C543G/G657E显著减弱了SMAD3的乙酰化(Figure 2I)。
质谱分析发现SMAD3 MH1区域赖氨酸20(K20)和赖氨酸117(K117)发生乙酰化,同时,这两个残基在不同物种间高度保守(Figure 2J)。在TGF-β1刺激下,赖氨酸到精氨酸的突变降低KAT6A诱导的SMAD3乙酰化,而赖氨酸和精氨酸的同时突变消除SMAD3的乙酰化(Figure 2K)。使用纯化的重组活性KAT6A和重组WT SMAD3或K20R或K117R突变体进行体外KAT实验证实KAT6A对K20和K117的乙酰化(Figure 2L)。此外,与WT SMAD3相比,乙酰化模拟K20/117Q突变体增强了SMAD3与KAT6A的关联,而非乙酰化K20/117R突变体抑制了它们的相互作用(Figure 2M)。
因此,这些数据表明,KAT6A可以直接乙酰化SMAD3的K20和K117残基。
三、SMAD3的K20/K117乙酰化上调免疫反应相关细胞因子
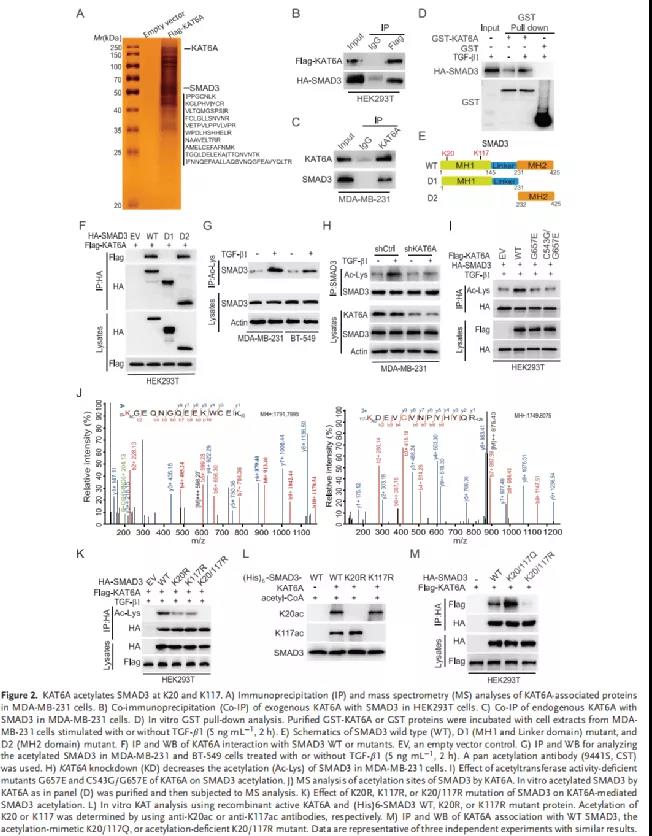
鉴于SMAD3是一个关键转录因子,作者假设SMAD3的K20/K117乙酰化增强了其转录活性。为验证这一假设,在MDA-MB-231/sgSMAD3细胞中进行了RNA-Seq,这些细胞过表达Flag-SMAD3 WT或精氨酸赖氨酸共同突变体(2KQ突变体)。测序分析出554个在2KQ突变体组高表达的基因(Figure 3A)。这554个基因在与炎症和免疫反应相关信号通路中高度富集(Figure 3B)。接下来,结合TCGA数据库,作者筛选出无转移和复发的TNBC患者样本中高表达的14个基因(Figure 3C)。基因集富集分析显示,KAT6A乙酰化的SMAD3(SMAD3-2KQ)上调炎症和免疫反应相关的(IL-6、IL-22和TNFα)信号通路(Figure 3D)。此外,QPCR分析发现,2KQ突变体促进了MDA-MB-231/sgSMAD3和BT-549/sgSMAD3细胞中IL-6、IL-22和TNFα的表达和分泌(Figure 3E-3F)。补充材料中,与SMAD3-WT相比,SMAD3-2KQ突变体显著提高了IL-6、IL-22和TNFA启动子的活性。ChIP-qPCR分析发现MDA-MB和BT-249细胞中SMAD3可与IL-6、IL-22和TNFA启动子结合。TGF-β1刺激确实可以以KAT6A依赖的方式导致这些细胞因子表达的增加。综上,SMAD3的K20/K117乙酰化上调免疫应答相关细胞因子的表达。四、SMAD3K的20/117位点乙酰化后能增加与KAT6A介导的TRIM24阅读蛋白的H3K23乙酰化,从而增强SMAD3的活性
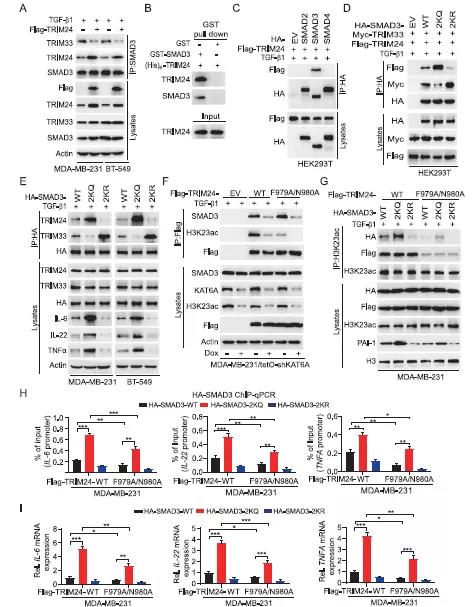
文献表明,TRIM24是TRIM超家族中TRIM33相关的辅助因子,也是KAT6A介导的H3K23ac在癌症中的阅读蛋白。因此作者研究了K20/117乙酰化是否能影响SMAD3与TRIM33和/或TRIM24的结合作用。结果发现,在TGF-β1刺激的MDA-MB-231和BT-549细胞中,TRIM24和TRIM33均与SMAD3结合(Figure 4A-B),且过表达TRIM24后,其与SMAD3的结合增强了,而TRIM33与SMAD3的结合却遭到了破坏(Figure 4A)。另外发现,除了SMAD3之外,SMAD2和SMAD4与TRIM24没有特异性关联(Figure 4C)。此外,通过补充材料说明TRIM24蛋白的中间段的393-823氨基酸残基介导了与SMAD3的结合。过表达TRIM33后发现SMAD3-TRIM24的相互结合作用减少。以上结果说明TRIM24和TRIM33竞争性与TGF-β1激活的SMAD3蛋白相结合。接下来,作者探索K20/117乙酰化对SMAD3与TRIM24和TRIM33结合的影响。与野生型SMAD3相比,乙酰化拟合2KQ突变体增强了TRIM24-SMAD3的相互作用,抑制了TRIM33与SMAD3的结合。而乙酰化缺失的2KQ突变体则得到了相反的结果(Figure 4D-4E)。2KQ突变体异位表达增加了IL-6、IL-22和TGF-β1的蛋白表达(Figure 4E)。以上结果表明,SMAD3的乙酰化促进了其与致癌基因TRIM24的相互作用,并降低了其与TRIM33肿瘤抑制因子的结合,从而增强了SMAD3的致癌活性。作者通过文献得到假设:KAT6A乙酰化H3K23招募TRIM24-SMAD3复合物,增强SMAD3-染色质相互作用,进而导致乳腺癌中SMAD3活化增强。首先,作者采用TRIM24 F979A/N980A的突变体破坏了TRIM24-H3K23ac的交互作用,但对TRIM24与SMAD3的相关性没有影响(Figure 4F)。在MDA-MB-231细胞中,共表达HA-tagged的SMAD3 WT、2KQ或2KR突变体与WT TRIM24 F979A/N980A突变体。结果得到,与WT TRIM24相比,F979A/N980A突变降低H3K23ac与TRIM24以及SMAD3 WT和突变体的关联(Figure 4G),也降低了SMAD3 WT和2KQ突变体与IL-6、IL-22和TNFα启动子的结合作用, 从而抑制IL-6、IL-22和TNF-β1的表达(Figure 4H-4I)。与SMAD3 WT与F979A/N980A突变共转染组相比,2KQ突变显著增加其与IL-6、IL-22和TNFA启动子的结合以及增强IL-6、IL-22和TNFA的表达(Figure 4H-I)。这些结果表明SMAD3 2KQ突变增加IL-6、IL-22和TNFA的表达与H3K23ac没有相关性,但H3K23ac结合能进一步增加SMAD3 2KQ突变体的活性。综上,KAT6A乙酰化SMAD3促进了SMAD3与H3K23ac阅读器TRIM24的结合,KAT6A介导的H3K23ac进一步增加了TRIM24/SMAD3-染色质关联,从而导致SMAD3信号激活增强。五、SMAD3的K20/K117乙酰化通过增强CSC和MDSCs招募促进TNBC的转移
以上结果提示,KAT6A/SMAD3轴可能影响TNBC中的乳腺癌干细胞样细胞(BCSC)特性。dox诱导KAT6A的敲降降低了IL-6、IL-22和TNFα 的蛋白表达、p-STAT3、CSC相关蛋白SOX2和CD44的表达、ALDH +亚种群、细胞成球能力(Figure 5A-5D)。以上实验结果能被拟乙酰化SMAD3的异位表达的2KQ突变体所逆转(Figure 5A-5D)。此外,与对照组相比,KAT6A KD增加了E-cadherin的表达,降低了N-cadherin、ZEB1、Slug和Snail的表达,但在拟乙酰化SMAD3的异位表达的2KQ突变体组中未观察到此变化趋势(补充材料)。综上,KAT6A-SMAD3轴能促进TNBC中的BCSC特性。为了研究KAT6A依赖SMAD3乙酰化对TNBC转移的调控作用,作者在有或没有KAT6A KD的4T1/sgSmad3细胞中表达Flag-SMAD3 WT和2KR突变体,从IL-6,IL-22,TNFα的表达、细胞增殖、迁移、侵袭、MDSCs招募、肺的微小转移等方面说明(补充材料)KAT6A诱导的SMAD3乙酰化能促进TNBC的转移。接下来,作者利用4T1小鼠乳腺癌转移模型,研究SMAD3乙酰化对体内MDSCs招募的影响。与未诱导细胞相比,dox诱导的KAT6A KD降低了IL-6、IL-22和TNFα、p-STAT3的表达(Figure 5E),KAT6A KD抑制肿瘤生长、肺转移,延长小鼠存活期(Figure 5F-5J),抑制MDSCs向转移性肺组织和原位瘤的募集以及转移性肺组织CD4+/CD8+ T细胞耗竭(Figure 5F-5M+补充材料)。以上结果均能被乙酰化模拟SMAD3 2KQ突变体所逆转(Figure 5E-5M)。故,KAT6A依赖SMAD3乙酰化通过MDSCs招募促进TNBC转移。为了进一步证明乳腺癌细胞分泌的促炎因子在SMAD3介导的肿瘤转移中的关键作用,作者构建了IL-6 KO 4T1细胞并发现IL-6 KO抑制了SMAD3-2KQ诱导的MDSCs募集(补充材料)。并在体内SMAD3-2KQ-诱导的MDSCs招募实验中也得到了证实(补充实验)。以上结果说明促炎因子IL-6亦参与SMAD3介导的肿瘤转移中过程。综上,KAT6A诱导的SMAD3乙酰化不仅增强乳腺癌干细胞样细胞特性,同时通过持续产生免疫应答相关细胞因子诱导MDSCs招募,促进乳腺癌的进展。<section sty

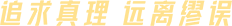


 2022-02-28 15:34:03
2022-02-28 15:34:03






 湘公网安备
湘公网安备